The oldest frog-line fossils are easiest to misunderstand when they are treated as prototypes of the modern leap. The name Salientia points toward jumping, and the living anuran body plan is so dominated by launch mechanics that it is tempting to read the whole lineage backward from a single explosive movement. Triadobatrachus massinoti, from the Early Triassic of Madagascar, is useful because it resists that shortcut.[1][2] It already sits on the frog side of the amphibian tree, but it does not yet look like an animal built around efficient saltation.
That is the real lineage lesson in 2026. The frog body plan was assembled in parts. The trunk shortened and the pelvis began to reorganize before the full jump-specialist package took hold.[1][2][5] Triadobatrachus preserves that unevenness unusually well: the specimen combines a still-long presacral region, discrete caudal vertebrae, and a less consolidated sacro-caudal complex with hindlimb and pelvic traits that point toward later frog geometry.[1][2] The result is not a failed frog. It is a stem salientian that shows how far the lineage had moved before high-performance jumping became the default reading.
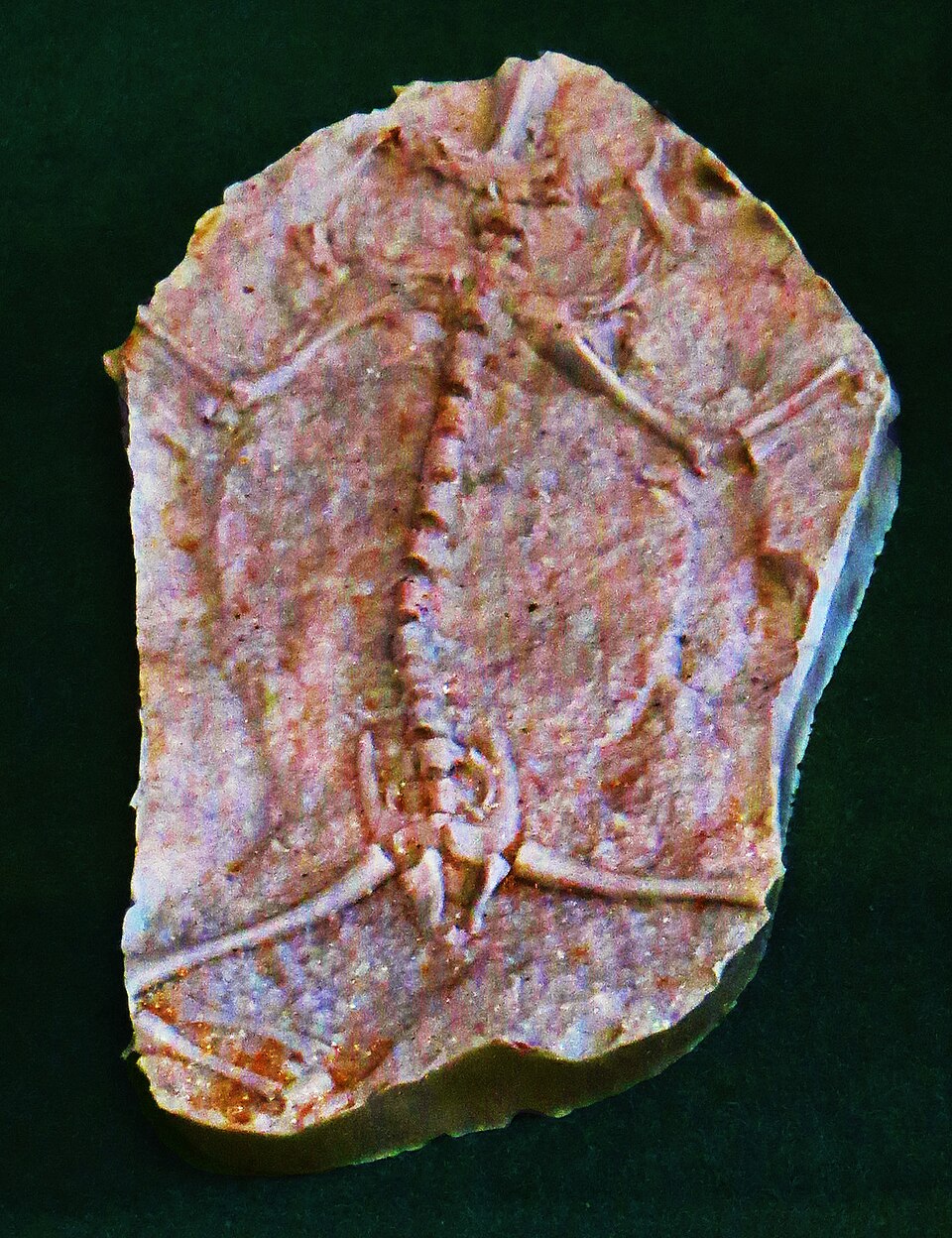
Image context: the cover uses a real museum photograph of the Triadobatrachus specimen from Wikimedia Commons. It belongs here because this article is about whole-body sequence. One photographed fossil does more work than a dramatic reconstruction: the long back and tail remain plainly visible even as the pelvic region starts to read as frog-like.[6]
1) The earliest salientian still kept more trunk than a modern frog could afford
Ascarrunz and colleagues' 2016 redescription remains the cleanest anatomical reset.[1] Their micro-CT-based re-examination updated the vertebral formula of Triadobatrachus to 15 presacrals, 1 sacral, and 10 or 11 caudals.[1] That count matters because it keeps the animal outside the compact axial design that makes modern frogs feel instantly recognizable. Living frogs usually operate with a far shorter presacral column and with the tail transformed into a urostyle rather than a string of separate caudal vertebrae.[2]
In other words, the earliest well-known salientian had already started moving away from the generalized tetrapod condition, but had not yet crossed into the rigid, shortened trunk of crown anurans.[1][2] The old habit of calling Triadobatrachus "the first frog" is therefore both useful and dangerous. Useful, because it belongs on the frog total-group side of the split. Dangerous, because it invites readers to imagine a modern jump mechanic in an animal that still carried too much back and tail to behave like a finished launcher.[1][2]
That long middle section is not a trivial leftover. It is the reason the fossil matters. If the trunk remained relatively extended while other features were already shifting, then the anuran body plan did not appear as one synchronized invention. The lineage was being edited module by module.[1][5]
2) Limb proportions point to walking first, not to full saltation
The locomotor argument becomes sharper in Lires, Soto, and Gomez's 2016 analysis.[2] Instead of assuming that every early frog-like feature must have served jumping from the outset, they tested Triadobatrachus in an explicit comparative framework using limb measurements from frogs, salamanders, and lizards.[2] Their conclusion is the one worth keeping close: the main locomotor mode of T. massinoti was best matched by salamander-like lateral undulatory movement, not by specialized jumping.[2]
That conclusion is valuable because it changes the causal order. The classic frog package contains elongated hindlimbs, reorganized pelvic mechanics, and a shortened, stiffened body that can transmit force cleanly during launch.[2][3] Triadobatrachus had only part of that package. Some postcranial features later useful for jumping were already present, but Lires and colleagues argued that these features were more plausibly exaptations at this stage, traits that predated the fully specialized behavior they later helped support.[2]
This keeps the article anchored in a broader paleontological rule: a trait can belong to the ancestry of a later function without having evolved for that function in its first appearance. That is exactly why the fossil matters for lineage context rather than for mascot value. It gives frog evolution a staggered timetable.[2][5]
3) By the Early Jurassic, the jump machine had become much more coherent
The contrast with Prosalirus shows what changed next.[3] In their 1995 Nature paper, Shubin and Jenkins described an Early Jurassic frog that already carried hallmarks of a far more consolidated anuran launch system: fusion of the caudal vertebrae into a urostyle, a rod-like ilium, and elongate hind limbs.[3] Their framing was explicit. The evolution of saltation in frogs involved a caudopelvic reorganization that could transmit hindlimb force into the axial skeleton during a jump.[3]
That is why Triadobatrachus is best read as a before-picture rather than a weak version of the same design. Between the Early Triassic stem salientian and the Early Jurassic frog, the body becomes more compact, the tail is re-engineered, and the pelvis becomes more fully committed to launch mechanics.[1][2][3] The jump does not stand at the beginning of the story. It stands at the point where several earlier anatomical tendencies finally lock together.
The consequence is simple but important. Frog evolution should not be narrated as "legs lengthened, therefore jumping began." The anatomy had to become integrated first. A long trunk with separate tail vertebrae and incomplete sacro-caudal consolidation belongs to a different locomotor world from a frog with a urostyle and a more force-transmitting pelvis.[1][2][3]
4) The lineage context is now broader than one Malagasy specimen
The broader stem-frog record supports that staged reading. Evans and Borsuk-Bialynicka's Early Triassic Czatkobatrachus from Poland already showed that salientians were not a one-locality curiosity and that some lineages could look slightly more derived than Triadobatrachus in parts of the vertebral column and elbow region.[4] That matters because it turns early frog evolution into a distributed experiment across Pangaea rather than a single fossil anecdote.[3][4]
More recent synthesis work keeps pressing in the same direction. Perez-Ben, Lires, and Gomez argue that the old habit of treating jumping as the root explanation for the entire anuran Bauplan no longer holds cleanly once limb data from extant and fossil frogs are analyzed together in phylogenetic context.[5] Their point is not that jumping became unimportant. It is that the origin of the frog body plan has to be separated from the later ecological and mechanical diversification of frogs as jumpers, walkers, swimmers, burrowers, and climbers.[5]
That is the secure way to read Triadobatrachus now. It preserves a frog-line animal in which the body has already moved away from the ancestral tetrapod pattern, yet still stops short of the full launch-ready architecture that later anurans would refine.[1][2][3][5] The fossil therefore does not show a primitive jump champion. It shows a lineage under construction.
The boundary matters as much as the claim. Triadobatrachus does not tell us that no early salientian could ever hop or lunge. It does not settle every step between Triassic stem frogs and Jurassic crownward forms. What it does secure is narrower and better: frogs became frog-like before they became efficient jump specialists, and the route to the modern leap ran through a longer sequence of trunk reduction, pelvic reorganization, and behavioral reinterpretation than the old slogan allowed.[1][2][5]
Sources
- Eduardo Ascarrunz, Jean-Claude Rage, Pierre Legreneur, and Michel Laurin, "Triadobatrachus massinoti, the earliest known lissamphibian (Vertebrata: Tetrapoda) re-examined by μCT scan, and the evolution of trunk length in batrachians," Contributions to Zoology 85(2) (2016) - redescription of the specimen, updated vertebral formula, and discussion of early batrachian trunk reduction.
- Andres I. Lires, Ignacio M. Soto, and Raul O. Gomez, "Walk before you jump: New insights on early frog locomotion from the oldest known salientian," Paleobiology 42(4) (2016) - comparative limb analysis arguing that Triadobatrachus is best matched by salamander-like lateral undulatory locomotion rather than specialized jumping.
- Neil H. Shubin and Farish A. Jenkins Jr., "An Early Jurassic jumping frog," Nature 377 (1995) - description of Prosalirus and the caudopelvic features associated with a more integrated jumping mechanism.
- Susan E. Evans and Magdalena Borsuk-Bialynicka, "A stem-group frog from the Early Triassic of Poland," Acta Palaeontologica Polonica 43(4) (1998) - Early Triassic stem-frog evidence from Poland showing that salientian diversification was broader than one Malagasy specimen.
- Celeste M. Perez-Ben, Andres I. Lires, and Raul O. Gomez, "Frog limbs in deep time: is jumping locomotion at the roots of the anuran Bauplan?" Paleobiology 50(1) (2024 issue; published online 2023) - synthesis testing whether the classic jumping-origin explanation for frog anatomy still holds across fossil and extant limb data.
- Wikimedia Commons file page for the museum photograph of the Triadobatrachus specimen used as the lead image.