最早那批蛙线化石,最容易被误读的时刻,正是它们被拿来充当现代跳跃机器原型的时刻。Salientia 这个名字本身就带着“跳跃”的方向感,而现生无尾类的身体方案,又几乎被起跳这一动作彻底支配,于是人们很自然地把整条谱系倒着读回去。来自马达加斯加早三叠世的 Triadobatrachus massinoti,真正重要的地方,恰好在于它挡住了这条省事的路径。[1][2] 它已经站在蛙类这一侧,却还没有成为一具围绕高效弹射来组织自身的身体。
这正是 2026 年最值得保留的谱系判断。蛙类身体方案并非一次到位的发明,它经历的是一段分层完成的组装过程。躯干先缩短,骨盆先开始改写,整套高效率跳跃系统则在后面才逐步锁定。[1][2][5] Triadobatrachus 之所以格外有用,就在于它把这种参差保留得很清楚:这件标本一面仍有偏长的前荐椎区、分离的尾椎,以及尚未完全整合的荐尾复合体,另一面又已经出现若干指向后世蛙类几何关系的骨盆与后肢特征。[1][2] 它更像一只把蛙类身体如何分段成形这件事直接摊开的基干显尾类动物。
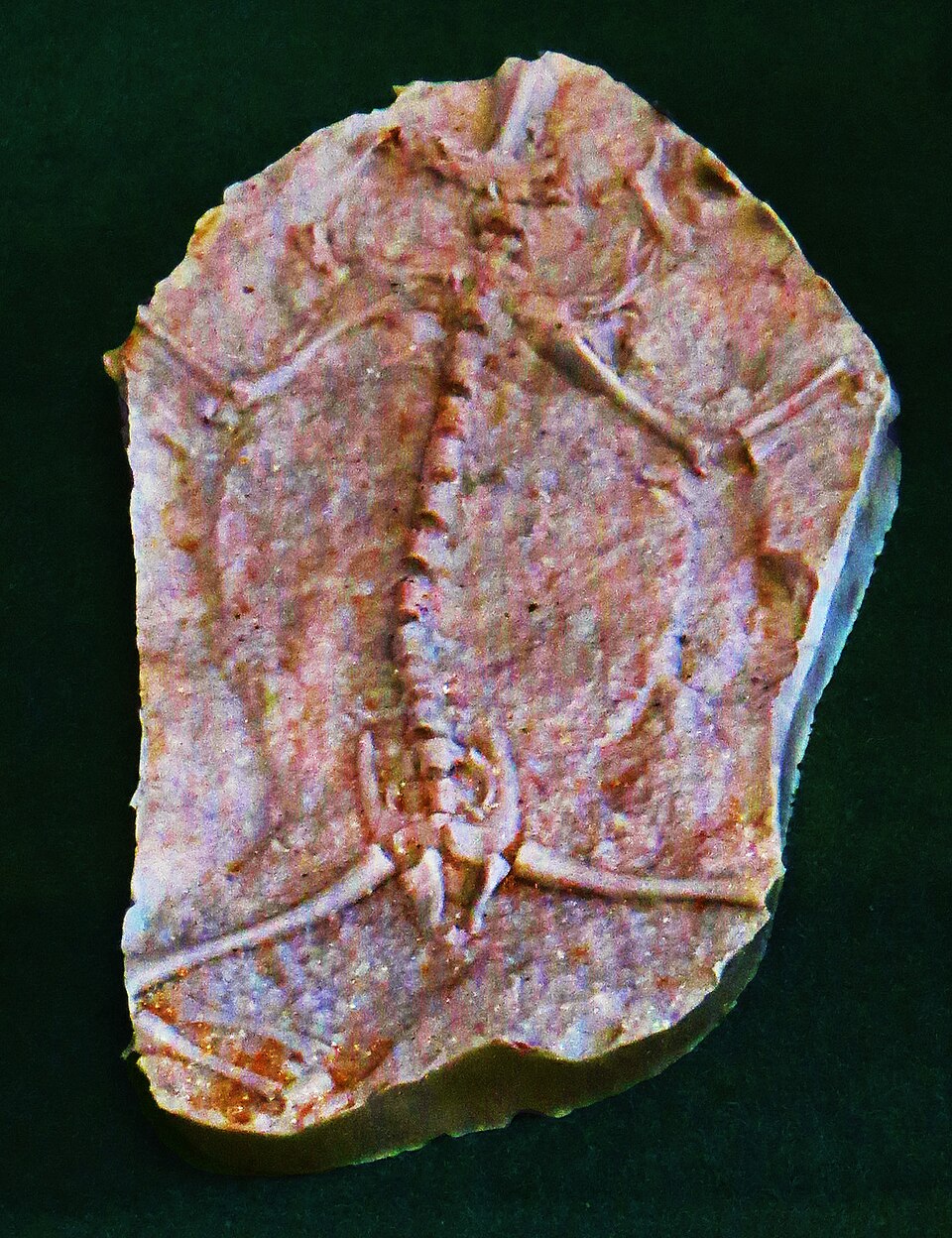
配图说明:题图采用 Wikimedia Commons 上公开的 Triadobatrachus 博物馆照片。它适合这篇文章,因为本文真正关心的是整具身体的次序关系。面对一件真实化石,较长的背部与尾部仍然清楚可见,骨盆区域又已经开始显露蛙类的轮廓,这比任何戏剧化复原图都更能把问题讲明白。[6]
1)最早的显尾类,依旧保留着现代青蛙负担不起的躯干
Ascarrunz 等人在 2016 年的重描述,至今仍是这件标本较稳的解剖学重置。[1] 这篇基于微型 CT 的研究,把 Triadobatrachus 的椎骨公式更新为 15 枚荐前椎、1 枚荐椎,以及 10 或 11 枚尾椎。[1] 这个数字之所以重要,在于它把这只动物稳稳留在现代无尾类那套紧凑轴向设计之外。现生青蛙通常拥有更短的荐前椎区,尾部也已不再是一串分离的尾椎,而是收束为一根尾杆式的 urostyle。[2]
也就是说,这只目前最重要的早期显尾类,已经明显偏离了广义四足动物的原始身体,却还没有跨进冠群无尾类那种短而紧的躯干结构。[1][2] 因此,把 Triadobatrachus 叫作“第一只青蛙”既有帮助,也容易带偏阅读。它有帮助,是因为它确实站在蛙类总群这一边;它容易带偏,是因为这个说法太容易让人把一套现代跳跃身体硬套回去,而这具化石显然仍然背着过长的脊背与尾部,不足以像成熟的发射器那样工作。[1][2]
这段较长的身体并非无关紧要的残留。它本身就是化石的意义所在。若躯干仍然延长,而其他模块已经开始改写,那就说明无尾类身体方案并非整套一起出现的,整条谱系是在一块一块地重写自身。[1][5]
2)四肢比例指向的首先是行走,而并非完整跳跃
进入运动方式这一层,Lires、Soto 与 Gomez 在 2016 年的分析把问题收得更紧。[2] 他们没有把每一项早期蛙类特征都直接解释成“原本就是为了跳跃”,而是把 Triadobatrachus 放进一套明确的比较框架中,与青蛙、蝾螈和蜥蜴的肢骨比例一起分析。[2] 最需要记住的结论,是他们对 T. massinoti 主要运动方式的判断:它最接近的是 类似蝾螈的侧向波动式行走,距离专门化跳跃仍有相当距离。[2]
这一点之所以关键,在于它改写了因果顺序。经典无尾类身体方案包含较长的后肢、被重新组织的骨盆力学,以及能够把力量干净传递出去的短而较硬的躯干。[2][3] Triadobatrachus 只拥有其中一部分。若干后来对跳跃十分重要的后躯特征,当时已经出现,Lires 等人却指出,这些结构在这一阶段更像 后起的功能借用基础,也就是先存在,随后才被跳跃系统重新利用的特征。[2]
这让整篇文章的判断回到一个更宽的古生物学原则上:一个特征可以处在后来功能的祖先链条里,却不等于它在首次出现时,就已经是为那项功能而存在。Triadobatrachus 正因为如此,更适合被放在演化脉络里阅读,而并非被当作某种“跳跃传奇的开场 mascot”。它把蛙类身体成形的时序,拆成了更可信的几段。[2][5]
3)到了早侏罗世,跳跃机器才开始真正收束成形
把 Triadobatrachus 与 Prosalirus 放在一起,对比会更清楚。[3] Shubin 与 Jenkins 在 1995 年那篇 Nature 论文里,描述了一只来自早侏罗世的蛙类,它已经具备一套明显更收束的无尾类跳跃结构:尾椎融合形成 urostyle、髂骨呈杆状、后肢延长。[3] 他们写得很直接,蛙类盐跃的演化,需要一套经过重写的尾骨盆复合体,把后肢力量传入中轴骨骼,再把起跳变成可重复的力学方案。[3]
这也解释了为何 Triadobatrachus 最适合被读成一张“前景图”,而并非同一种系统的薄弱版本。从早三叠世的基干显尾类,到早侏罗世的真正蛙类,身体变得更紧凑,尾部被重新工程化,骨盆也更彻底地投入了起跳机制。[1][2][3] 跳跃并不站在故事的开端,它站在多条较早的解剖趋势终于锁合起来的位置上。
顺着这个变化去看,蛙类演化就不该再被写成“后肢变长,于是开始跳”。真正发生的,是整套身体先被整合。一具带着较长躯干、分离尾椎、荐尾区尚未充分收束的身体,和一只拥有 urostyle、能够更有效传力的骨盆系统的青蛙,属于两个完全不同的运动世界。[1][2][3]
4)谱系背景早已超出马达加斯加这一件标本
更宽的干群记录也支持这种分段读法。Evans 与 Borsuk-Bialynicka 对波兰早三叠世 Czatkobatrachus 的研究说明,显尾类从来并非单一产地里的孤立奇观,而且某些支系在椎骨与肘部区域已经比 Triadobatrachus 稍显进步。[4] 这一层意义很重要,因为它把早期蛙类演化从“某件马达加斯加化石的单点故事”,扩展成泛盘古大陆上的分布式试验。[3][4]
更新近的综合研究则把这一点说得更直接。Perez-Ben、Lires 与 Gomez 指出,一旦把化石与现生青蛙的四肢数据放进系统发育框架里共同分析,过去那种把“跳跃”当作整套无尾类身体起源总解释的旧习惯,已经不再稳固。[5] 他们并非说跳跃不重要,而是把两件原本总被粘在一起的事情拆开:一件是蛙类身体方案如何起源,另一件则是青蛙后来如何在跳跃、步行、游泳、掘穴与攀爬之间持续分化。[5]
这就是今天阅读 Triadobatrachus 较稳的位置。它保存了一只已经明显脱离祖先型四足动物模式、却仍然停在完整发射式身体之前的蛙线动物。[1][2][3][5] 这件化石展示的,是一条仍在施工中的谱系。
边界也需要保留下来。Triadobatrachus 没有把任何早期显尾类的轻跃、扑跃或短距冲动行为全部排除在外,它也没有把从三叠纪干群蛙到侏罗纪更进步类型之间的每一步都一次说尽。它真正稳稳支撑起来的判断更窄,也更值得信赖:蛙类是在成为高效率跳跃专家之前,先变得越来越像蛙;通向现代一跃的道路,中间经历了更长的一段躯干缩短、骨盆重组与功能重释的过程。[1][2][5]
来源
- Eduardo Ascarrunz、Jean-Claude Rage、Pierre Legreneur、Michel Laurin,"Triadobatrachus massinoti, the earliest known lissamphibian (Vertebrata: Tetrapoda) re-examined by μCT scan, and the evolution of trunk length in batrachians," Contributions to Zoology 85(2)(2016)——用于标本重描述、更新后的椎骨公式,以及早期蛙蝾共同支躯干缩短的讨论。
- Andres I. Lires、Ignacio M. Soto、Raul O. Gomez,"Walk before you jump: New insights on early frog locomotion from the oldest known salientian," Paleobiology 42(4)(2016)——基于四肢比例的比较分析,主张 Triadobatrachus 的主要运动方式更接近蝾螈式侧向波动,而并非专门化跳跃。
- Neil H. Shubin、Farish A. Jenkins Jr.,"An Early Jurassic jumping frog," Nature 377(1995)——用于 Prosalirus 的描述,以及与成熟跳跃相关的尾骨盆结构整合。
- Susan E. Evans、Magdalena Borsuk-Bialynicka,"A stem-group frog from the Early Triassic of Poland," Acta Palaeontologica Polonica 43(4)(1998)——用于波兰早三叠世干群蛙证据,以及显尾类早期分布并不局限于马达加斯加这一点。
- Celeste M. Perez-Ben、Andres I. Lires、Raul O. Gomez,"Frog limbs in deep time: is jumping locomotion at the roots of the anuran Bauplan?" Paleobiology 50(1)(2024 期次,2023 年先行上线)——综合化石与现生无尾类四肢数据,重新检验“跳跃是否位于蛙类身体方案根部”这一老问题。
- 本文题图所用 Triadobatrachus 博物馆照片的 Wikimedia Commons 文件页。