Hurdia victoria is a useful fossil precisely because it began as the wrong kind of object. The name was attached in 1912 to an isolated triangular carapace from the Burgess Shale, not to a complete animal with all its parts conveniently attached.[3][4] For decades, other pieces of the same biological puzzle sat under different labels: lateral carapaces, frontal appendages, and mouth structures that had been compared with other Burgess animals or treated as parts of uncertain affinity.[1][3] The animal became clear only when paleontologists stopped asking what one dramatic plate was and started asking which recurring fragments belonged together.
That makes Hurdia a better close-reading subject than a simple predator profile. It is not just another Cambrian creature with a strange silhouette. It is a case study in how a soft-bodied animal can be disassembled by death, burial, collection, and early taxonomy, then reassembled by patient comparison across specimens. The fossil's central lesson is methodological: in Burgess Shale paleontology, a named part may be only one word in a sentence that took a century to read.
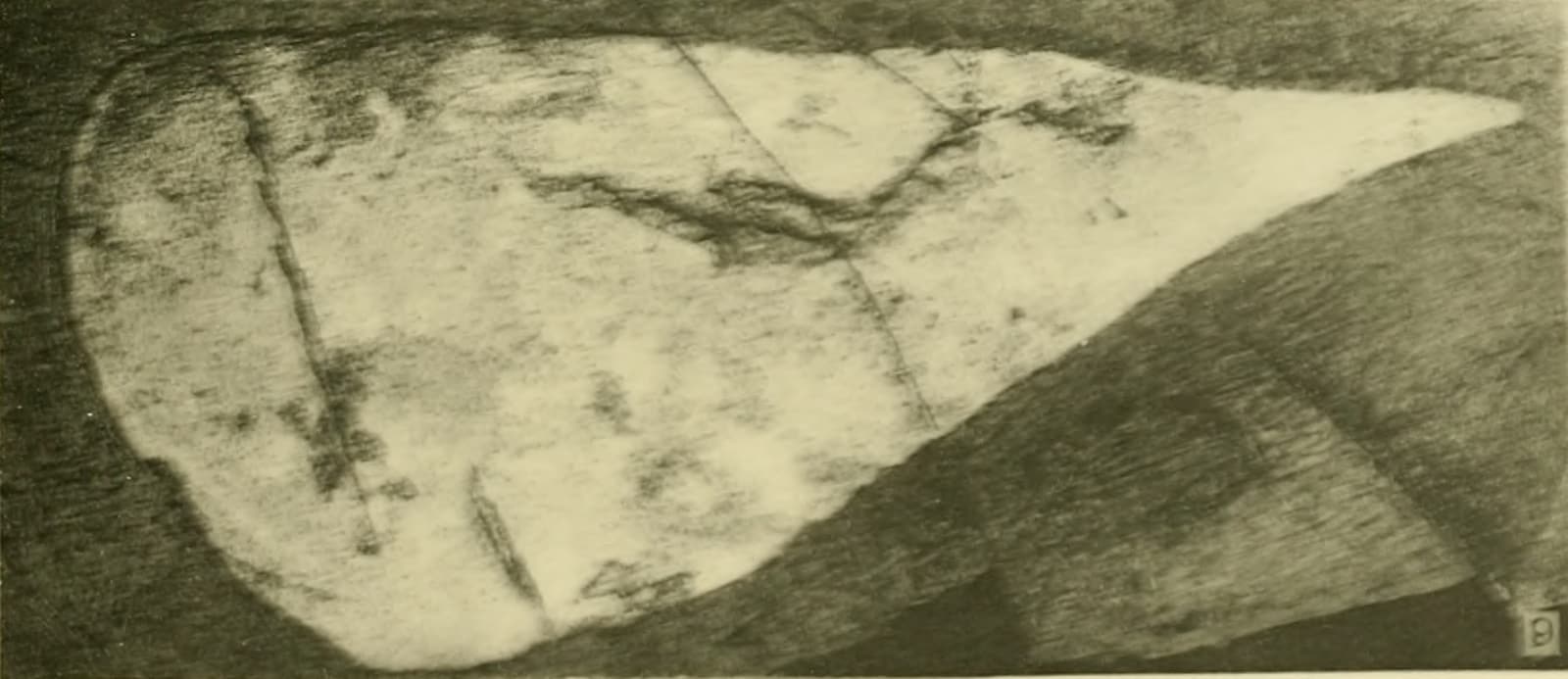
Image context: the cover uses the Wikimedia Commons file for the Hurdia victoria holotype, USNM PAL 57718, originally figured from Walcott's 1912 work and now associated with the Smithsonian National Museum of Natural History.[4] The photograph keeps the article anchored to a real archival fossil surface rather than a life reconstruction.
The named fossil was a carapace, not the animal
The holotype is visually modest: a dark, triangular piece on shale, preserved with enough outline to be recognizable but not enough anatomy to disclose an entire body plan.[4] That modesty is the point. Walcott's original Hurdia was a part fossil. The Royal Ontario Museum's Burgess Shale account identifies the structure as the central carapace, or H-element, of the head complex.[3] Alone, it could not tell a reader where the eyes sat, how the mouth worked, how the body swam, or whether the animal belonged near ordinary crustaceans, trilobites, or something farther down the arthropod stem.
This is why Hurdia resists the quick museum-label habit. A large front plate makes a strong visual claim, but it can also hide the animal's real organization. If the central carapace is treated as the whole story, Hurdia becomes a triangular mystery. If it is treated as one head sclerite in a larger apparatus, the fossil opens into a more precise radiodont problem.[2][3]
The 2009 Science paper by Daley, Budd, Caron, Edgecombe, and Collins was the decisive reset. It tied the old Hurdia carapace to a larger anomalocaridid animal and placed that animal in the early euarthropod stem-group conversation.[1] The important move was not only naming a complete organism. It was matching anatomical parts that had been historically scattered across separate taxonomic stories.
The puzzle pieces had separate histories
The ROM history of research makes the old confusion unusually clear. The carapace called Hurdia was one part. Proboscicaris, first described as a different kind of carapace, became another part of the same front complex. Frontal appendages had their own history, including early association with Sidneyia and later treatment as "Appendage F" of uncertain affinity. Mouthparts had also been tangled with the older Peytoia problem, one of the classic Burgess Shale cases in which isolated anatomy was misread before whole-body specimens clarified the association.[3]
Read that way, Hurdia is not embarrassing evidence that early paleontologists were careless. It is evidence that the fossil record was giving them an unusually difficult task. Radiodont bodies were not mineralized skeletons that fell apart along familiar vertebrate or trilobite lines. They were soft-bodied animals with sclerites, flaps, appendages, and mouthparts that could preserve separately. A quarry full of parts can look like a quarry full of animals until enough repeated associations show which pieces travel together.[1][2][3]
The 2013 morphology and systematics paper made that assembly work deeper. Daley, Budd, and Caron examined hundreds of specimens from British Columbia and Utah, using a larger sample to refine Hurdia morphology and systematics rather than relying on one spectacular body.[2] That scale matters. A single complete fossil can be persuasive, but a population of complete and disarticulated remains can show which variation is taxonomic, which is taphonomic, and which is just the normal scatter of anatomy after decay.
The head was a three-part shield around a predator's toolkit
Once assembled, the head of Hurdia stops being a single triangle. The ROM description divides the anterior region into a complex of non-biomineralized carapaces: one dorsal triangular H-element and two lateral paddle-shaped P-elements, arranged around the front of the head.[3] This is the key correction to the cover image. The photographed H-element is real and historically central, but it is not a portrait. It is one roof piece from a more elaborate head structure.
Under that shield was the feeding machinery. ROM describes a ventral mouth made of a circlet of overlapping plates, plus frontal appendages with segmented construction and elongated inner spines.[3] Those appendages are why Hurdia belongs with radiodonts rather than with ordinary shell-bearing arthropods. The animal was not biting with jaws in the modern arthropod sense. It was using paired frontal limbs and a circular mouth apparatus in a body plan that sits outside the crown-group arthropod template.[1][3]
That distinction matters because Hurdia should not be flattened into "Cambrian shrimp" or "Cambrian monster." Its anatomy is more disciplined than either phrase. It had radiodont hallmarks: frontal appendages, a circular mouth, lateral swimming flaps, and head sclerites that framed the anterior body.[1][3] But the large carapace complex gives it a different emphasis from the more familiar public image of Anomalocaris. It is a predator whose first visual signal is armor-like architecture, not just grasping arms.
The body turns the carapace into a life habit
The rear half of the animal keeps the close reading honest. ROM describes a posterior body with seven to nine segments extending into triangular flaps, associated with elongated blade-like structures interpreted as gills, and ending in rounded lobes.[3] Complete specimens are reported up to about 20 centimeters, while disarticulated fragments suggest animals reaching roughly 30 centimeters.[3] Those numbers matter less as trivia than as a scale correction. Hurdia was large for its world, but it was not a mythic sea monster. It was a mobile Cambrian animal whose ecological role has to be inferred from a working body.
The best current ecological reading is bounded. ROM lists the life habit as mobile and nektobenthic, with a predatory feeding strategy.[3] The large dorsal carapace, upward-facing eyes, stubby body, and appendage form have supported interpretations of an animal spending much of its time near the seafloor, possibly feeding on prey associated with the substrate.[3] That is a stronger image than a generic open-water hunter. It puts the shield, eyes, appendages, and flaps into one behavioral setting.
Still, the carapace itself should not be over-solved. ROM notes that the function of the frontal carapace remains unknown, even though feeding-related roles have been suggested.[3] That uncertainty is valuable. The fossil gives enough anatomy to rule out many old isolated-part interpretations, but not enough to turn every surface into a settled function. A good close reading has to keep both facts alive: Hurdia is far clearer than it was in 1912, and some of its most conspicuous anatomy still resists a single tidy purpose.
Why Hurdia still matters
The strongest claim for Hurdia is not that it is the strangest radiodont. The Burgess Shale already has plenty of candidates for that kind of attention. Its deeper value is that it shows how scientific understanding can change when a fossil is moved from object to association. A triangle becomes an H-element. A supposed separate carapace becomes a lateral plate. A loose appendage becomes one member of a frontal pair. A mouthpart joins a head and swimming body. The animal appears only after the parts stop competing as isolated names.[1][2][3]
That reading also makes Hurdia useful for the wider arthropod-origin story. Radiodonts sit near the stem of euarthropod evolution, carrying some features that point toward later arthropod organization without already being crown-group arthropods.[1][2] Hurdia therefore matters twice: as a reconstructed animal in its own Cambrian ecology, and as a reminder that early arthropod evolution was not a smooth ladder of familiar forms. It was a field of body plans whose parts had to be sorted before their evolutionary meaning could be argued.
The holotype image is almost austere after that. It does not show the whole predator. It shows the starting error and the starting evidence at the same time: a single triangular fossil, meaningful but incomplete. Read alone, it is a named shard. Read through a century of associated specimens, it becomes the roof of a Cambrian animal that finally came into focus when paleontology learned how to put the fragments back together.
Sources
- Allison C. Daley, Graham E. Budd, Jean-Bernard Caron, Gregory D. Edgecombe, and Desmond Collins, "The Burgess Shale Anomalocaridid Hurdia and Its Significance for Early Euarthropod Evolution," Science 323, no. 5921 (2009), bibliographic DOI record.
- Allison C. Daley, Graham E. Budd, and Jean-Bernard Caron, "Morphology and systematics of the anomalocaridid arthropod Hurdia from the Middle Cambrian of British Columbia and Utah," Journal of Systematic Palaeontology 11, no. 7 (2013), Crossref metadata record.
- Royal Ontario Museum, "Hurdia victoria," Burgess Shale fossil page - specimen history, morphology, abundance, ecology, and references.
- Wikimedia Commons, "File:Hurdia victoria USNM PAL 57718.jpg" - source page for the archival holotype photograph used as the lead image.